Chemiosmosi, forza motrice protonica e fosforilazione ossidativa
In ogni trasferimento di un elettrone attraverso l’ETS, l’elettrone perde energia, ma con alcuni trasferimenti, l’energia viene immagazzinata come energia potenziale usandola per pompare ioni idrogeno (H+) attraverso una membrana. Nelle cellule procariotiche, gli H+ sono pompati all’esterno della membrana citoplasmatica (chiamata spazio periplasmatico nei batteri gram-negativi e gram-positivi), e nelle cellule eucariotiche, sono pompati dalla matrice mitocondriale attraverso la membrana mitocondriale interna nello spazio intermembrana. C’è una distribuzione non uniforme di H+ attraverso la membrana che stabilisce un gradiente elettrochimico perché gli ioni H+ sono caricati positivamente (elettrici) e c’è una concentrazione maggiore (chimica) su un lato della membrana. Questo gradiente elettrochimico formato dall’accumulo di H+ (noto anche come protone) su un lato della membrana rispetto all’altro è chiamato forza motrice protonica (PMF). Poiché gli ioni coinvolti sono H+, si stabilisce anche un gradiente di pH, con il lato della membrana che ha una maggiore concentrazione di H+ che è più acido. Oltre all’uso della PMF per produrre ATP, come discusso in questo capitolo, la PMF può anche essere usata per guidare altri processi energeticamente sfavorevoli, tra cui il trasporto dei nutrienti e la rotazione dei flagelli per la motilità.
L’energia potenziale di questo gradiente elettrochimico generato dall’ETS causa la diffusione dell’H+ attraverso una membrana (la membrana plasmatica nelle cellule procariotiche e la membrana interna dei mitocondri nelle cellule eucariotiche). Questo flusso di ioni idrogeno attraverso la membrana, chiamato chemiosmosi, deve avvenire attraverso un canale nella membrana attraverso un complesso enzimatico legato alla membrana chiamato ATP sintasi (Figura \(\PageIndex{1}). La tendenza al movimento in questo modo è molto simile all’acqua accumulata su un lato di una diga, che si muove attraverso la diga quando viene aperta. L’ATP sintasi (come una combinazione della presa e del generatore di una diga idroelettrica) è una proteina complessa che agisce come un piccolo generatore, girando con la forza degli H+ che diffondono attraverso l’enzima, lungo il loro gradiente elettrochimico da dove ci sono molti H+ che si respingono a vicenda a dove ci sono meno H+. Nelle cellule procariotiche, l’H+ scorre dall’esterno della membrana citoplasmatica nel citoplasma, mentre nei mitocondri eucarioti, l’H+ scorre dallo spazio intermembrana alla matrice mitocondriale. La rotazione delle parti di questa macchina molecolare rigenera l’ATP dall’ADP e dal fosfato inorganico (Pi) attraverso la fosforilazione ossidativa, un secondo meccanismo per produrre ATP che raccoglie l’energia potenziale immagazzinata in un gradiente elettrochimico.
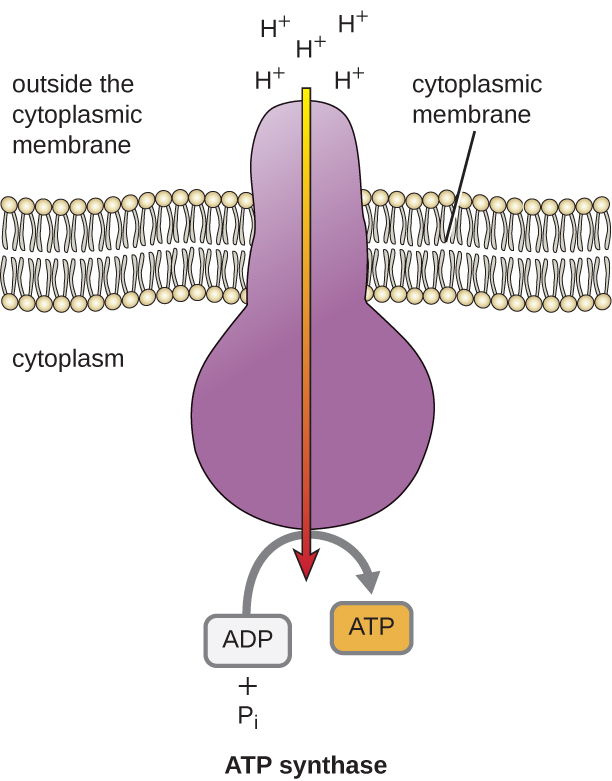
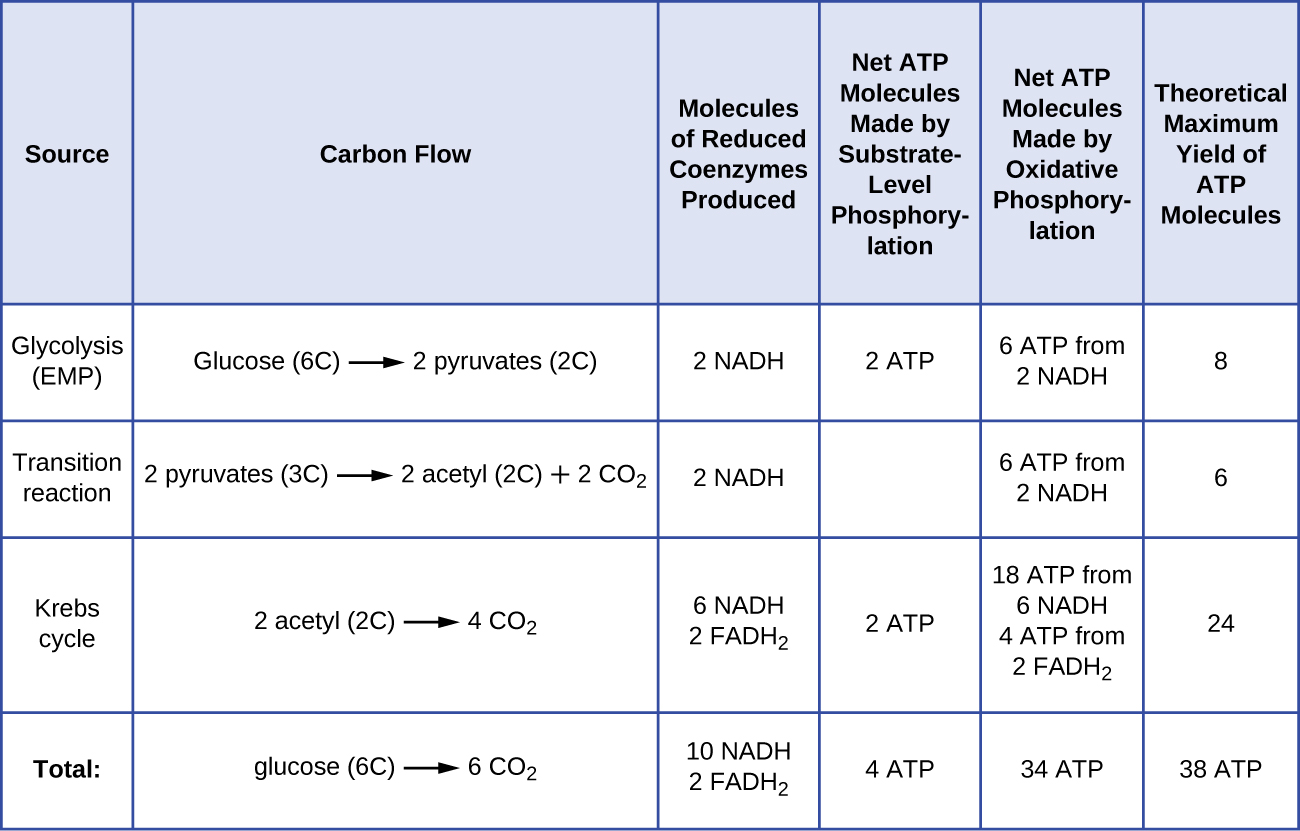
Il numero di molecole di ATP generate dal catabolismo del glucosio varia. Per esempio, il numero di ioni idrogeno che i complessi del sistema di trasporto degli elettroni possono pompare attraverso la membrana varia tra le diverse specie di organismi. Nella respirazione aerobica nei mitocondri, il passaggio di elettroni da una molecola di NADH genera una forza motrice protonica sufficiente per fare tre molecole di ATP per fosforilazione ossidativa, mentre il passaggio di elettroni da una molecola di FADH2 genera una forza motrice protonica sufficiente per fare solo due molecole di ATP. Così, le 10 molecole di NADH prodotte per glucosio durante la glicolisi, la reazione di transizione e il ciclo di Krebs portano abbastanza energia per produrre 30 molecole di ATP, mentre le due molecole di FADH2 prodotte per glucosio durante questi processi forniscono abbastanza energia per produrre quattro molecole di ATP. Complessivamente, il rendimento massimo teorico di ATP prodotto durante la respirazione aerobica completa del glucosio è di 38 molecole, di cui quattro sono prodotte dalla fosforilazione a livello del substrato e 34 dalla fosforilazione ossidativa (Figura \(\PageIndex{2}). In realtà, la resa totale di ATP è di solito inferiore, da una a 34 molecole di ATP, a seconda che la cellula usi la respirazione aerobica o anaerobica; nelle cellule eucariotiche, una parte dell’energia viene spesa per trasportare gli intermedi dal citoplasma ai mitocondri, influenzando la resa di ATP.
La figura \(\PageIndex{2}}) riassume i rendimenti massimi teorici di ATP da vari processi durante la respirazione aerobica completa di una molecola di glucosio.
Esercizio \(\PageIndex{1})
Quali sono le funzioni della forza motrice protonica?