Chemiosmosis, protonernas drivkraft och oxidativ fosforylering
Vid varje överföring av en elektron genom ETS förlorar elektronen energi, men vid vissa överföringar lagras energin som potentiell energi genom att den används för att pumpa vätejoner (H+) genom ett membran. I prokaryota celler pumpas H+ till utsidan av cytoplasmimembranet (kallat periplasmautrymmet i gramnegativa och grampositiva bakterier), och i eukaryota celler pumpas de från mitokondrialmatrisen över det inre mitokondrialmembranet till intermembranutrymmet. Det finns en ojämn fördelning av H+ över membranet som skapar en elektrokemisk gradient eftersom H+-joner är positivt laddade (elektriskt) och det finns en högre koncentration (kemiskt) på ena sidan av membranet. Denna elektrokemiska gradient som bildas av ansamlingen av H+ (även kallad proton) på ena sidan av membranet jämfört med den andra kallas för protonmotivkraft (PMF). Eftersom de inblandade jonerna är H+ bildas också en pH-gradient, där den sida av membranet som har den högre koncentrationen av H+ är surare. Förutom användningen av PMF för att tillverka ATP, som diskuteras i detta kapitel, kan PMF också användas för att driva andra energimässigt ogynnsamma processer, inklusive näringstransport och flagellas rotation för motilitet.
Den potentiella energin i denna elektrokemiska gradient som genereras av ETS gör att H+ diffunderar över ett membran (plasmamembranet i prokaryota celler och det inre membranet i mitokondrier i eukaryota celler). Detta flöde av vätejoner över membranet, som kallas kemosmos, måste ske genom en kanal i membranet via ett membranbundet enzymkomplex som kallas ATP-syntas (figur \(\(\PageIndex{1}\)). Tendensen till rörelse på detta sätt är ungefär som att vatten som ackumuleras på ena sidan av en damm rör sig genom dammen när den öppnas. ATP-syntas (som en kombination av intag och generator i en vattenkraftsdamm) är ett komplext protein som fungerar som en liten generator, som vrider sig med hjälp av kraften från de H+ som diffunderar genom enzymet, nerför deras elektrokemiska gradient från där det finns många ömsesidigt avstötande H+ till där det finns färre H+. I prokaryota celler flödar H+ från utsidan av cytoplasmimembranet in i cytoplasman, medan H+ i eukaryota mitokondrier flödar från intermembranutrymmet till mitokondriematrisen. När delarna i detta molekylära maskineri vänder på varandra återskapas ATP från ADP och oorganiskt fosfat (Pi) genom oxidativ fosforylering, en andra mekanism för att tillverka ATP som skördar den potentiella energi som finns lagrad i en elektrokemisk gradient.
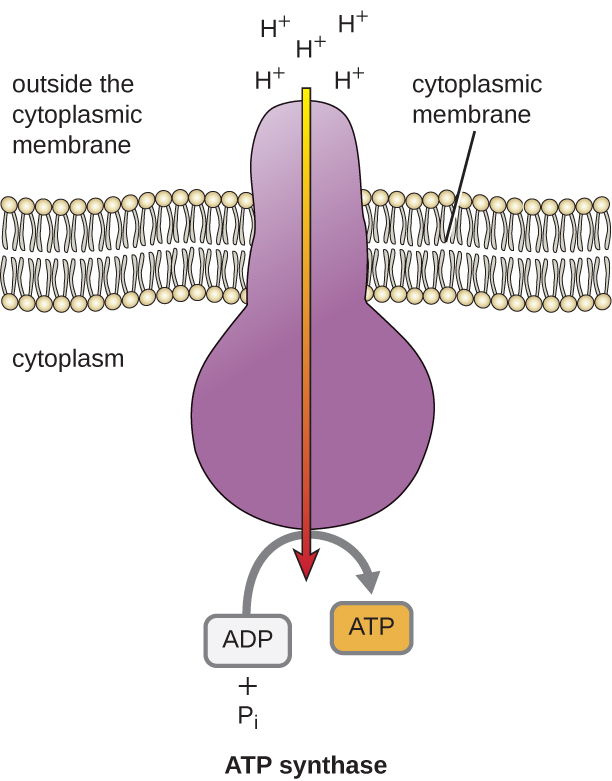
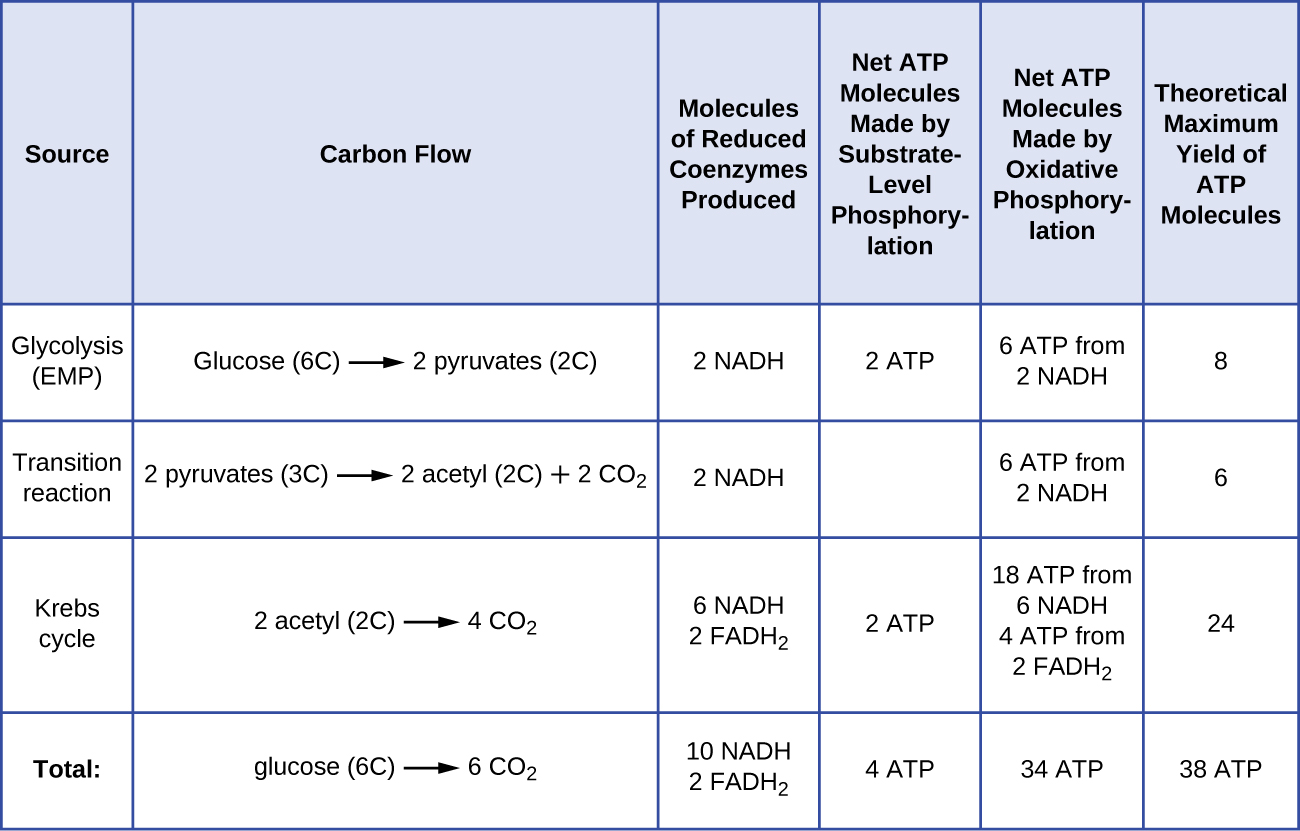
Antalet ATP-molekyler som genereras från katabolismen av glukos varierar. Till exempel varierar antalet vätejoner som elektrontransportsystemkomplexen kan pumpa genom membranet mellan olika arter av organismer. Vid aerob andning i mitokondrier genererar passagen av elektroner från en molekyl NADH tillräckligt med protonmotivationskraft för att skapa tre ATP-molekyler genom oxidativ fosforylering, medan passagen av elektroner från en molekyl FADH2 genererar tillräckligt med protonmotivationskraft för att skapa endast två ATP-molekyler. De tio NADH-molekyler som bildas per glukos under glykolysen, övergångsreaktionen och Krebscykeln medför således tillräckligt med energi för att bilda 30 ATP-molekyler, medan de två FADH2-molekyler som bildas per glukos under dessa processer ger tillräckligt med energi för att bilda fyra ATP-molekyler. Totalt sett är det teoretiska maximala utbytet av ATP under en fullständig aerob andning av glukos 38 molekyler, varav fyra genom fosforylering på substratnivå och 34 genom oxidativ fosforylering (figur \(\PageIndex{2}\)). I verkligheten är det totala ATP-utbytet vanligtvis mindre, från en till 34 ATP-molekyler, beroende på om cellen använder aerob respiration eller anaerob respiration. I eukaryota celler går en del energi åt för att transportera intermediärer från cytoplasman till mitokondrierna, vilket påverkar ATP-utbytet.
Figur \(\PageIndex{2}\) sammanfattar det teoretiska maximala utbytet av ATP från olika processer under fullständig aerob respiration av en glukosmolekyl.
Övningsuppgift \(\PageIndex{1}\)
Vilka funktioner har protonmotivkraften?