Chemiosmosis, protonmotivationskraft og oxidativ fosforylering
I hver overførsel af en elektron gennem ETS mister elektronen energi, men ved nogle overførsler gemmes energien som potentiel energi ved at bruge den til at pumpe hydrogenioner (H+) gennem en membran. I prokaryote celler pumpes H+ til ydersiden af cytoplasmamembranen (kaldet det periplasmatiske rum i gramnegative og grampositive bakterier), og i eukaryote celler pumpes de fra mitokondrialmatrixen over den indre mitokondriemembran ind i intermembranrummet. Der er en ujævn fordeling af H+ på tværs af membranen, som etablerer en elektrokemisk gradient, fordi H+-ioner er positivt ladede (elektrisk), og der er en højere koncentration (kemisk) på den ene side af membranen. Denne elektrokemiske gradient, der dannes af ophobningen af H+ (også kendt som en proton) på den ene side af membranen sammenlignet med den anden, kaldes protonmotivationskraften (PMF). Da de involverede ioner er H+, opstår der også en pH-gradient, idet den side af membranen, der har den højere koncentration af H+, er mere sur. Ud over brugen af PMF til at fremstille ATP, som omtalt i dette kapitel, kan PMF også bruges til at drive andre energimæssigt ugunstige processer, herunder næringsstoftransport og flagelrotation med henblik på motilitet.
Den potentielle energi i denne elektrokemiske gradient, der genereres af ETS, får H+ til at diffundere over en membran (plasmamembranen i prokaryote celler og den indre membran i mitokondrier i eukaryote celler). Denne strøm af hydrogenioner gennem membranen, kaldet kemosmose, skal ske gennem en kanal i membranen via et membranbundet enzymkompleks kaldet ATP-syntase (figur \(\(\PageIndex{1}\)). Tendensen til bevægelse på denne måde svarer meget til, at vand, der ophobes på den ene side af en dæmning, bevæger sig gennem dæmningen, når den åbnes. ATP-syntase (som en kombination af indtag og generator i en hydroelektrisk dæmning) er et komplekst protein, der fungerer som en lille generator, der drejer ved hjælp af kraften fra de H+, der diffunderer gennem enzymet, ned ad deres elektrokemiske gradient fra et sted, hvor der er mange gensidigt frastødende H+ til et sted, hvor der er færre H+. I prokaryote celler strømmer H+ fra ydersiden af cytoplasmamembranen ind i cytoplasmaet, mens H+ i eukaryote mitokondrier strømmer fra intermembranrummet til mitokondriens matrix. Drejningen af delene i denne molekylære maskine regenererer ATP fra ADP og uorganisk fosfat (Pi) ved oxidativ fosforylering, en anden mekanisme til fremstilling af ATP, der høster den potentielle energi, der er lagret i en elektrokemisk gradient.
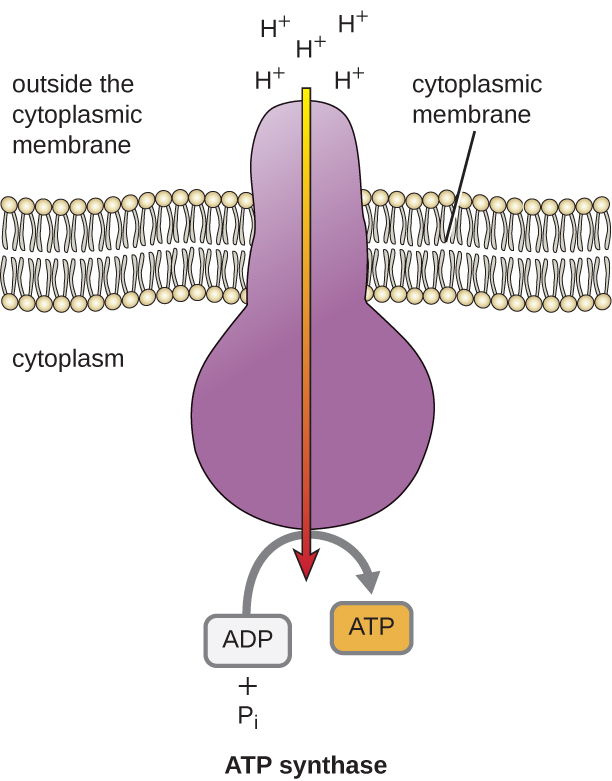
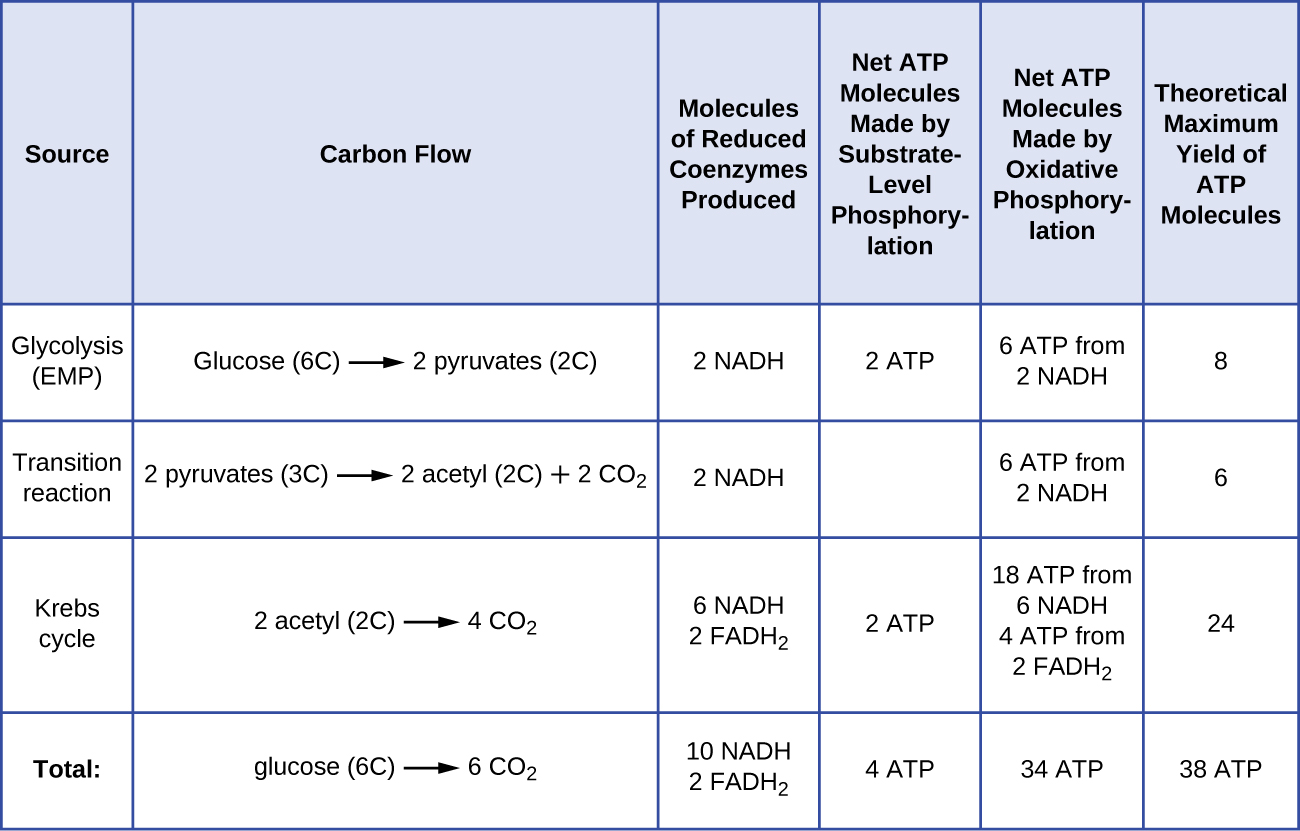
Antallet af ATP-molekyler, der dannes ved katabolisme af glukose, varierer. For eksempel varierer antallet af hydrogenioner, som elektrontransportsystemkomplekserne kan pumpe gennem membranen, mellem forskellige arter af organismer. Ved aerob respiration i mitokondrier genererer passage af elektroner fra ét molekyle NADH tilstrækkelig protonmotivationskraft til at danne tre ATP-molekyler ved oxidativ fosforylering, hvorimod passage af elektroner fra ét molekyle FADH2 genererer tilstrækkelig protonmotivationskraft til kun at danne to ATP-molekyler. De 10 NADH-molekyler, der dannes pr. glukose under glykolysen, overgangsreaktionen og Krebs-cyklussen, bærer således energi nok til at danne 30 ATP-molekyler, mens de to FADH2-molekyler, der dannes pr. glukose under disse processer, giver energi nok til at danne fire ATP-molekyler. Samlet set er det teoretiske maksimale udbytte af ATP fremstillet under den fuldstændige aerobe respiration af glukose 38 molekyler, hvoraf fire fremstilles ved fosforylering på substratniveau og 34 ved oxidativ fosforylering (figur \(\PageIndex{2}\)). I virkeligheden er det samlede ATP-udbytte normalt mindre, nemlig fra et til 34 ATP-molekyler, afhængigt af om cellen anvender aerob respiration eller anaerob respiration; i eukaryote celler bruges der en del energi på at transportere mellemprodukter fra cytoplasmaet til mitokondrierne, hvilket påvirker ATP-udbyttet.
Figur \(\PageIndex{2}\) opsummerer det teoretiske maksimale udbytte af ATP fra forskellige processer under den fuldstændige aerobe respiration af et glukosemolekyle.
Ovelse \(\PageIndex{1}\)
Hvad er funktionerne af den protonmotive kraft?