Quimiosmosis, Fuerza Motriz de Protones, y Fosforilación Oxidativa
En cada transferencia de un electrón a través del ETS, el electrón pierde energía, pero con algunas transferencias, la energía se almacena como energía potencial utilizándola para bombear iones de hidrógeno (H+) a través de una membrana. En las células procariotas, los H+ se bombean hacia el exterior de la membrana citoplasmática (llamada espacio periplásmico en las bacterias gramnegativas y grampositivas), y en las células eucariotas, se bombean desde la matriz mitocondrial a través de la membrana mitocondrial interna hacia el espacio intermembrana. Hay una distribución desigual de H+ a través de la membrana que establece un gradiente electroquímico porque los iones H+ tienen carga positiva (eléctrica) y hay una mayor concentración (química) en un lado de la membrana. Este gradiente electroquímico formado por la acumulación de H+ (también conocido como protón) en un lado de la membrana en comparación con el otro se denomina fuerza motriz de protones (PMF). Como los iones implicados son H+, también se establece un gradiente de pH, siendo más ácido el lado de la membrana que tiene una mayor concentración de H+. Más allá del uso de la PMF para producir ATP, como se discute en este capítulo, la PMF también puede utilizarse para impulsar otros procesos energéticamente desfavorables, incluyendo el transporte de nutrientes y la rotación de los flagelos para la motilidad.
La energía potencial de este gradiente electroquímico generado por la ETS hace que el H+ se difunda a través de una membrana (la membrana plasmática en las células procariotas y la membrana interna en las mitocondrias en las células eucariotas). Este flujo de iones de hidrógeno a través de la membrana, llamado quimiosmosis, debe ocurrir a través de un canal en la membrana a través de un complejo enzimático unido a la membrana llamado ATP sintasa (Figura \(\PageIndex{1}\)). La tendencia a moverse de esta manera es muy parecida al agua acumulada en un lado de una presa, que se mueve a través de la presa cuando se abre. La ATP sintasa (como una combinación de la toma y el generador de una presa hidroeléctrica) es una proteína compleja que actúa como un pequeño generador, girando por la fuerza de los H+ que se difunden a través de la enzima, por su gradiente electroquímico desde donde hay muchos H+ que se repelen mutuamente hasta donde hay menos H+. En las células procariotas, el H+ fluye desde el exterior de la membrana citoplasmática hacia el citoplasma, mientras que en las mitocondrias eucariotas, el H+ fluye desde el espacio intermembranal hacia la matriz mitocondrial. El giro de las piezas de esta máquina molecular regenera ATP a partir de ADP y fosfato inorgánico (Pi) mediante la fosforilación oxidativa, un segundo mecanismo de fabricación de ATP que aprovecha la energía potencial almacenada dentro de un gradiente electroquímico.
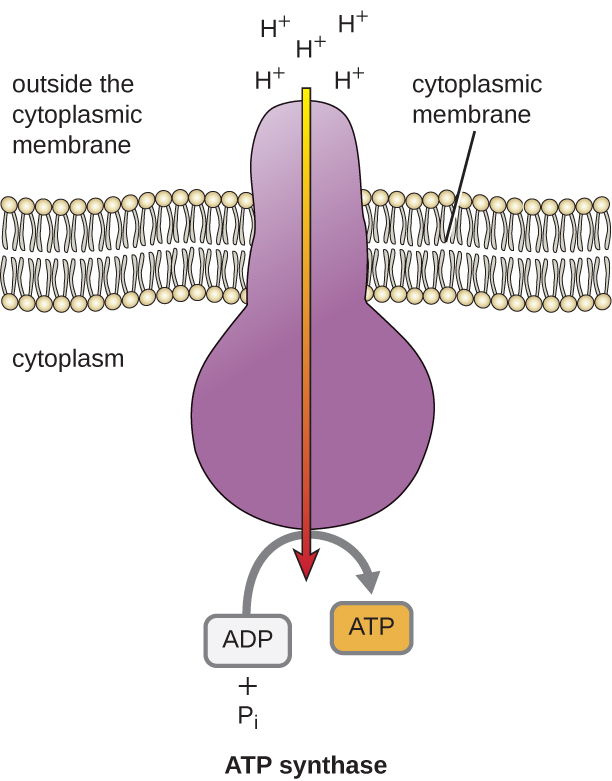
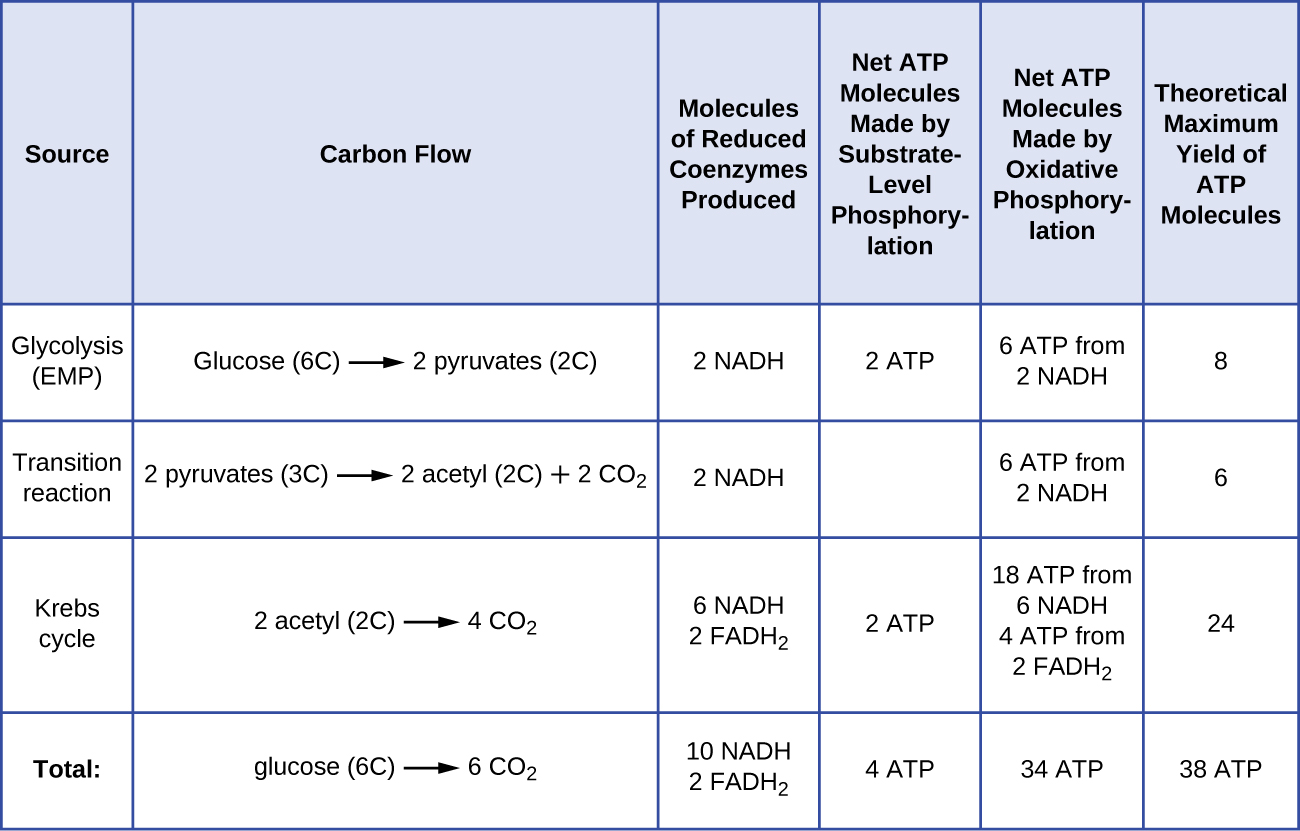
El número de moléculas de ATP generadas a partir del catabolismo de la glucosa varía. Por ejemplo, el número de iones de hidrógeno que los complejos del sistema de transporte de electrones pueden bombear a través de la membrana varía entre las diferentes especies de organismos. En la respiración aeróbica de las mitocondrias, el paso de electrones de una molécula de NADH genera suficiente fuerza motriz de protones para producir tres moléculas de ATP por fosforilación oxidativa, mientras que el paso de electrones de una molécula de FADH2 genera suficiente fuerza motriz de protones para producir sólo dos moléculas de ATP. Por lo tanto, las 10 moléculas de NADH producidas por la glucosa durante la glucólisis, la reacción de transición y el ciclo de Krebs llevan suficiente energía para producir 30 moléculas de ATP, mientras que las dos moléculas de FADH2 producidas por la glucosa durante estos procesos proporcionan suficiente energía para producir cuatro moléculas de ATP. En total, el rendimiento máximo teórico de ATP producido durante la respiración aeróbica completa de la glucosa es de 38 moléculas, de las cuales cuatro se producen por fosforilación a nivel de sustrato y 34 por fosforilación oxidativa (Figura \(\PageIndex{2}\)). En realidad, el rendimiento total de ATP suele ser menor, oscilando entre una y 34 moléculas de ATP, dependiendo de si la célula utiliza la respiración aeróbica o la respiración anaeróbica; en las células eucariotas, se gasta algo de energía para transportar los productos intermedios desde el citoplasma a la mitocondria, lo que afecta al rendimiento de ATP.
La figura \N(\PageIndex{2}\N) resume los rendimientos máximos teóricos de ATP de varios procesos durante la respiración aeróbica completa de una molécula de glucosa.
Ejercicio \(\PageIndex{1})
¿Cuáles son las funciones de la fuerza motriz de protones?