Hemiosmoosi, protonien liikkeellepanovoima ja oksidatiivinen fosforylaatio
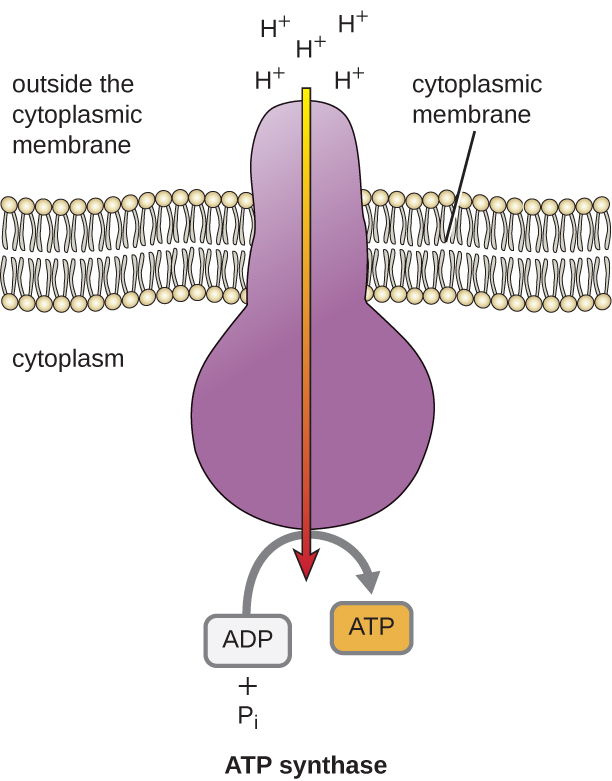
Kussakin elektronin siirrossa ETS:n läpi elektroni menettää energiaa, mutta joissakin siirroissa energia varastoituu potentiaalienergiaksi käyttämällä sitä vetyionien (H+) pumppaamiseen membraanin läpi. Prokaryoottisoluissa H+ pumpataan sytoplasmakalvon ulkopuolelle (jota kutsutaan periplasmaksi tilaksi gramnegatiivisissa ja grampositiivisissa bakteereissa), ja eukaryoottisoluissa ne pumpataan mitokondriomatriisista mitokondrioiden sisemmän kalvon läpi kalvojen väliseen tilaan. H+ jakautuu epätasaisesti kalvon poikki, mikä luo sähkökemiallisen gradientin, koska H+-ionit ovat positiivisesti varautuneita (sähköisesti) ja niiden pitoisuus on suurempi (kemiallisesti) kalvon toisella puolella. Tätä sähkökemiallista gradienttia, joka muodostuu H+ -ionien (tunnetaan myös nimellä protoni) kertymisestä kalvon toiselle puolelle verrattuna toiseen, kutsutaan protonien käyttövoimaksi (PMF). Koska kyseessä ovat H+ -ionit, syntyy myös pH-gradientti, jolloin kalvon se puoli, jolla on suurempi H+ -pitoisuus, on happamampi. Sen lisäksi, että PMF:ää käytetään ATP:n tuottamiseen, kuten tässä luvussa käsitellään, PMF:ää voidaan käyttää myös muiden energeettisesti epäedullisten prosessien käyttämiseen, kuten ravinteiden kuljetukseen ja lippulaivojen pyörimiseen liikkuvuutta varten.
Tämän ETS:n synnyttämän sähkökemiallisen gradientin potentiaalienergia saa aikaan sen, että H+ diffundoituu membraanin poikki (plasmamembraanin plasmakalvossa prokaryoottisoluissa ja sisäisen membraanin mitokondrioiden sisemmässä kalvossa eukaryoottisoluissa). Tämän kalvon läpi tapahtuvan vetyionien virtauksen, jota kutsutaan kemosmoosiksi, on tapahduttava kalvossa olevan kanavan kautta kalvoon sidotun ATP-syntaasiksi kutsutun entsyymikompleksin kautta (kuva \(\PageIndex{1}\)). Taipumus liikkua tällä tavoin on kuin padon toiselle puolelle kertynyt vesi, joka liikkuu padon läpi, kun se avataan. ATP-syntaasi (kuin vesivoimapadon imu- ja generaattorin yhdistelmä) on monimutkainen proteiini, joka toimii pienenä generaattorina, joka kääntyy entsyymin läpi diffundoituvien H+ -yhdisteiden voimasta alaspäin sähkökemiallisessa gradientissaan paikasta, jossa on paljon toisiaan hylkiviä H+ -yhdisteitä, paikkaan, jossa H+ -yhdisteitä on vähemmän. Prokaryoottisoluissa H+ virtaa sytoplasmakalvon ulkopuolelta sytoplasmaan, kun taas eukaryoottisissa mitokondrioissa H+ virtaa kalvojen välisestä tilasta mitokondriomatriisiin. Tämän molekyylikoneiston osien kääntäminen regeneroi ATP:tä ADP:stä ja epäorgaanisesta fosfaatista (Pi) oksidatiivisella fosforylaatiolla, joka on toinen ATP:n valmistusmekanismi, joka kerää sähkökemialliseen gradienttiin varastoitunutta potentiaalienergiaa.
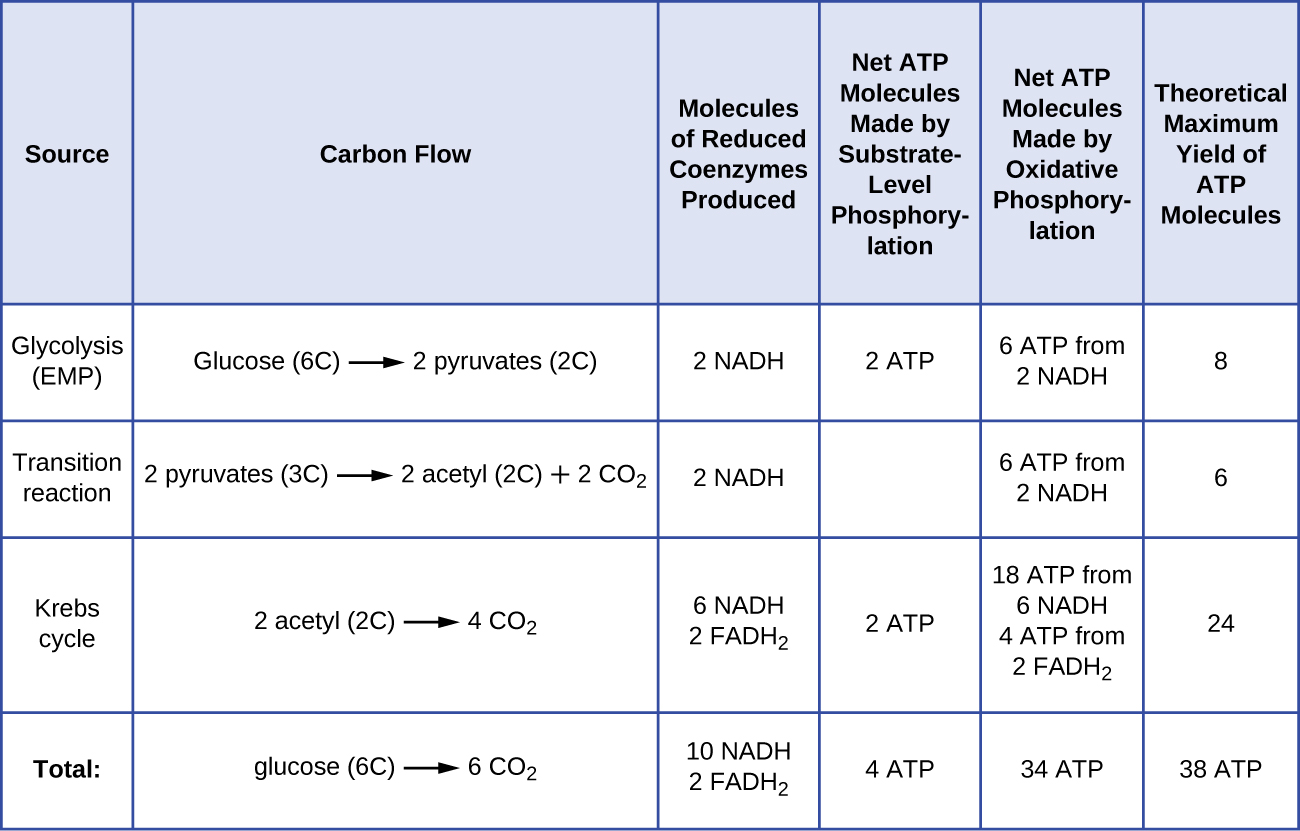
Glukoosin kataboliasta syntyvien ATP-molekyylien määrä vaihtelee. Esimerkiksi vetyionien määrä, jonka elektroninkuljetusjärjestelmäkompleksit voivat pumpata kalvon läpi, vaihtelee eri eliölajien välillä. Mitokondrioiden aerobisessa hengityksessä elektronien kulkeutuminen yhdestä NADH-molekyylistä tuottaa riittävästi protonien liikkeellepanevaa voimaa kolmen ATP-molekyylin valmistamiseen oksidatiivisella fosforylaatiolla, kun taas elektronien kulkeutuminen yhdestä FADH2-molekyylistä tuottaa riittävästi protonien liikkeellepanevaa voimaa vain kahden ATP-molekyylin valmistamiseen. Näin ollen glykolyysin, siirtymäreaktion ja Krebsin syklin aikana glukoosia kohti tuotetut 10 NADH-molekyyliä kuljettavat riittävästi energiaa 30 ATP-molekyylin tuottamiseen, kun taas näiden prosessien aikana glukoosia kohti tuotetut kaksi FADH2-molekyyliä tuottavat riittävästi energiaa neljän ATP-molekyylin tuottamiseen. Kaiken kaikkiaan glukoosin täydellisen aerobisen hengityksen aikana syntyvän ATP:n teoreettinen enimmäistuotto on 38 molekyyliä, joista neljä syntyy substraattitason fosforylaatiossa ja 34 oksidatiivisessa fosforylaatiossa (kuva \(\PageIndex{2}\)). Todellisuudessa ATP:n kokonaistuotanto on yleensä pienempi ja vaihtelee yhdestä 34 ATP-molekyylistä riippuen siitä, käyttääkö solu aerobista vai anaerobista hengitystä; eukaryoottisoluissa kuluu jonkin verran energiaa välituotteiden kuljettamiseen sytoplasmasta mitokondrioihin, mikä vaikuttaa ATP:n tuottoon.
Kuvassa \(\PageIndex{2}\) on yhteenveto ATP:n teoreettisista enimmäistuotoista eri prosesseista yhden glukoosimolekyylin täydellisen aerobisen hengityksen aikana.
Harjoitustehtävä \(\PageIndex{1}\)
Mitkä ovat protonimoottorivoiman tehtävät?