Chemiosmose, Proton Motive Force e Fosforilação Oxidativa
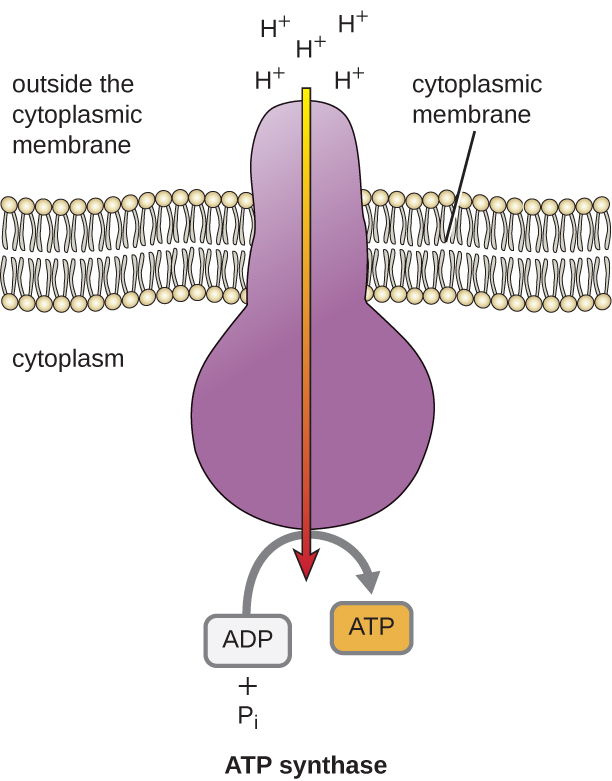
Em cada transferência de um elétron através da ETS, o elétron perde energia, mas com algumas transferências, a energia é armazenada como energia potencial, usando-a para bombear íons de hidrogênio (H+) através de uma membrana. Nas células procarióticas, o H+ é bombeado para o exterior da membrana citoplasmática (chamado espaço periplasmático em bactérias gram-negativas e gram-positivas), e nas células eucarióticas, são bombeados da matriz mitocondrial através da membrana mitocondrial interna para o espaço intermembrana. Há uma distribuição desigual de H+ através da membrana que estabelece um gradiente eletroquímico porque os íons H+ são positivamente carregados (elétricos) e há uma concentração maior (química) em um lado da membrana. Este gradiente eletroquímico formado pelo acúmulo de H+ (também conhecido como próton) em um lado da membrana em relação ao outro é chamado de força motriz do próton (PMF). Como os íons envolvidos são H+, também é estabelecido um gradiente de pH, sendo que o lado da membrana tem a maior concentração de H+ mais ácida. Além do uso do PMF para fazer ATP, como discutido neste capítulo, o PMF também pode ser usado para conduzir outros processos energeticamente desfavoráveis, incluindo transporte de nutrientes e rotação de flagelos para motilidade.
A energia potencial deste gradiente eletroquímico gerado pelo ETS faz com que o H+ se difunda através de uma membrana (a membrana plasmática em células procarióticas e a membrana interna em mitocôndrias em células eucarióticas). Este fluxo de íons de hidrogênio através da membrana, chamado quimiosmose, deve ocorrer através de um canal na membrana através de um complexo enzimático ligado à membrana chamado ATP synthase (Figura \PageIndex{1}}). A tendência para o movimento desta forma é muito semelhante à água acumulada num dos lados de uma barragem, movendo-se através da barragem quando aberta. A ATP synthase (como uma combinação da captação e gerador de uma barragem hidroelétrica) é uma proteína complexa que atua como um pequeno gerador, girando pela força do H+ que se difunde através da enzima, descendo seu gradiente eletroquímico de onde há muitos H+ que se repelem mutuamente para onde há menos H+. Nas células procarióticas, o H+ flui do exterior da membrana citoplasmática para o citoplasma, enquanto que nas mitocôndrias eucarióticas, o H+ flui do espaço intermembrana para a matriz mitocondrial. O giro das partes desta máquina molecular regenera ATP a partir do ADP e fosfato inorgânico (Pi) por fosforilação oxidativa, um segundo mecanismo para fazer ATP que colhe a energia potencial armazenada dentro de um gradiente eletroquímico.
>
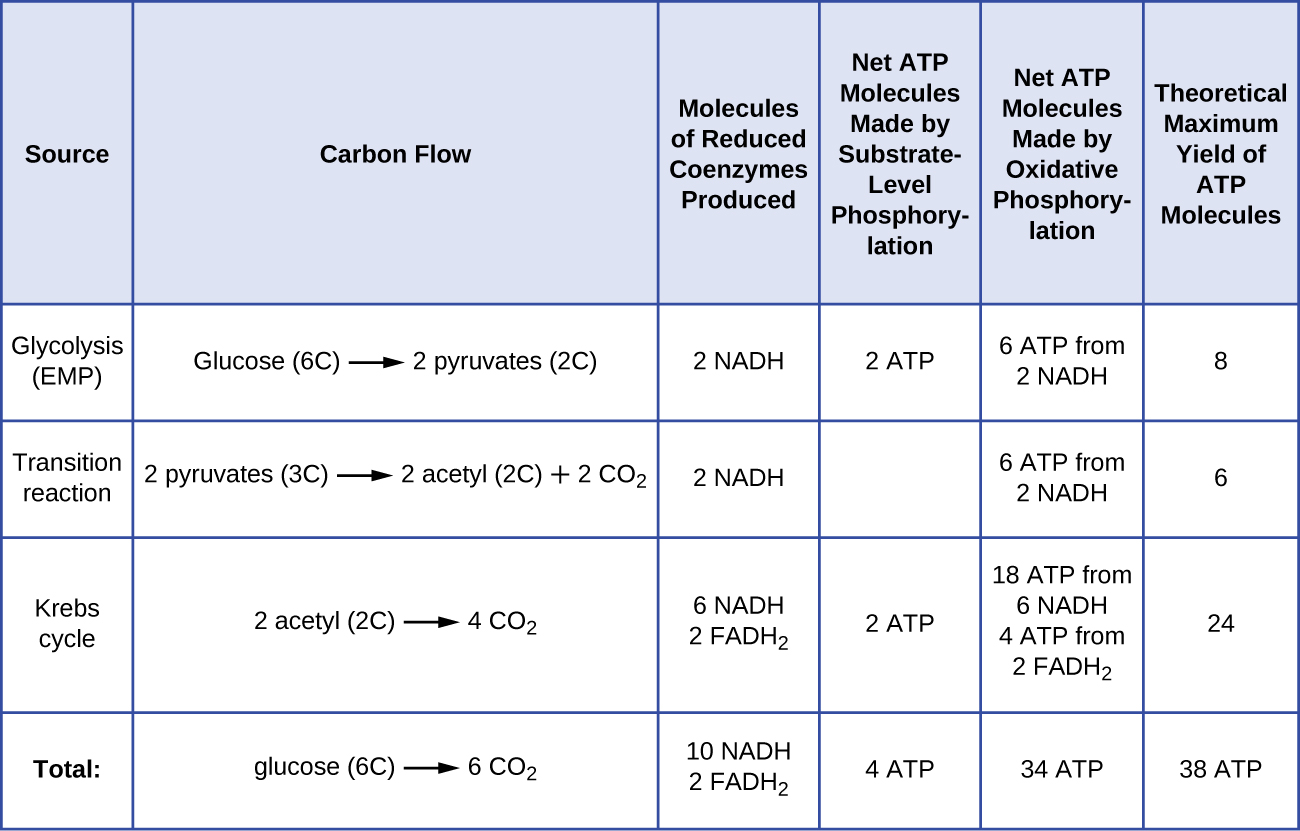
O número de moléculas de ATP geradas a partir do catabolismo da glicose varia. Por exemplo, o número de íons de hidrogênio que os complexos do sistema de transporte de elétrons podem bombear através da membrana varia entre as diferentes espécies de organismos. Na respiração aeróbica em mitocôndrias, a passagem de elétrons de uma molécula de NADH gera força motriz de prótons suficiente para fazer três moléculas de ATP por fosforilação oxidativa, enquanto a passagem de elétrons de uma molécula de FADH2 gera força motriz de prótons suficiente para fazer apenas duas moléculas de ATP. Assim, as 10 moléculas de NADH feitas por glicose durante a glicólise, a reação de transição e o ciclo de Krebs carregam energia suficiente para fazer 30 moléculas de ATP, enquanto as duas moléculas de FADH2 feitas por glicose durante esses processos fornecem energia suficiente para fazer quatro moléculas de ATP. Em geral, o rendimento máximo teórico de ATP feito durante a respiração aeróbica completa da glicose é de 38 moléculas, sendo quatro feitas por fosforilação ao nível do substrato e 34 feitas por fosforilação oxidativa (Figura \PageIndex{2}). Na realidade, a produção total de ATP é geralmente menor, variando de uma a 34 moléculas de ATP, dependendo se a célula está usando respiração aeróbica ou anaeróbica; em células eucarióticas, alguma energia é gasta para transportar intermediários do citoplasma para as mitocôndrias, afetando a produção de ATP.
Figure \(\PageIndex{2}) resume os rendimentos máximos teóricos de ATP de vários processos durante a respiração aeróbica completa de uma molécula de glucose.
Exercício \(\PageIndex{1}})
Quais são as funções da força motriz do próton?